Михаил Супотницкий (Россия)

Михаил Супотницкий (Россия) |
|
Феномен антигенного импринтинга в эпидемических, инфекционных и поствакцинальных процессах |
|
| Биопрепараты, 2014; (3):27–39 |
Супотницкий Михаил Васильевич (р. 1956) — кандидат
биологических наук, старший научный сотрудник, бывший военный микробиолог, полковник мед. службы
запаса. Автор многочисленных книг и публикаций по эпидемиологии возбудителей опасных инфекционных
болезней и проблемам биологической безопасности. Сайт М. Супотницкого — http://www.supotnitskiy.ru/avtor.htm
Оригинал здесь Феномен антигенного импринтинга (феномен первичного антигенного греха, анамнестический ответ) еще не стал объектом пристального исследования отечественных ученых. Цель работы — показать роль антигенного импринтинга в эпидемических, инфекционных и поствакцинальных процессах. Феномен заключается в том, что при повторном контакте иммунной системы с патогенным микроорганизмом или антигенами, включенными в состав вакцины, различия между прежним вариантом эпитопа антигена и его новым вариантом могут оказаться "не замеченными" иммунной системой, и она в течение всей жизни человека будет отвечать на тот антиген, который запомнила первым. Антигенный импринтинг развивается в отношении возбудителей инфекционных болезней, содержащих множество поверхностных эпитопов разной перекрестной специфичности, включающих доминантные эпитопы, перекрестно-реагирующие с антителами к антигенам таксономически малосвязанных возбудителей инфекционных болезней или имеющих незначительные отличия в пределах вида. Феномен наблюдается при взаимодействии иммунной системы с антигенно-дрейфующими микроорганизмами (ВИЧ, вирусы гриппа, лимфоцитарного хориоменингита) и микроорганизмами более стабильных видов, но циркулирующих конкурентно как множество разных серотипов и штаммов (лихорадки Денге, малярийные плазмодии и др.). Он предопределяет тяжесть течения болезни и структуру заболеваемость населения во время эпидемий (пандемий). Поствакцинальные осложнения, связанные с антигенным импринтингом, могут проявляться через десятилетия после проведения вакцинации. Вакцинация одной и той же вакциной может дать противоположные результаты в группах населения, имеющих разную эпидемическую историю и ранее многократно вакцинированных вакцинами различающегося антигенного состава. В настоящее время сформировалось по меньшей мере три направления конструирования вакцин, пытающихся устранить развитие этого феномена в ответ на инфекцию после проведенной вакцинации или повторной вакцинации: удаление перекрестно-реагирующих эпитопов у доминантного антигена возбудителя инфекции, активизация дендритных клеток специальными адъювантами, гетерологичный примирующий бустинг. В статье описан возможный алгоритм исследований антигенного импринтинга при доклиническом изучении вакцин. Библиографическое описание: Супотницкий М. В. Феномен антигенного импринтинга в инфекционных, эпидемических и поствакцинальных процессах. // Биопрепараты 2014; (3) с. 27–39. Antigen imprinting phenomenon (the phenomenon of original antigenic sin, anamnestic response) has not yet become a subject of intense research of national scientists. The purpose of the present study is to show its role in epidemic, infectious diseases and vaccine-related processes. The core of the phenomenon is during the repeated contact of the immune system with a pathogen or a vaccine, the differences between the old version of the antigenic epitope, and its new version could be "not detected" by the immune system and it will respond to the first to remember antigen during the life of an individual. Antigen imprinting develops in response to infectious agents, containing plenty of surface epitopes with various cross-specificity, including antigenic drifting microorganisms (HIV, influenza virus, lymphocytic choriomeningitis virus) and more stable species, but circulating competitively in the form of various serotypes and strains (Dengue fever, malaria parasites and other). It defines the severity of a disease and the structure of morbidity during epidemics (pandemics). Post-vaccination implications associated with antigen imprinting, occur decades after vaccination. Vaccination with the same vaccine may give different results in populations with different epidemiological background and previously repeatedly vaccinated by vaccines of different antigenic composition. There are at least three categories of vaccine designing, trying to eliminate the development of this phenomenon in response to infection after vaccination or revaccination: removing cross-reactive epitopes of an antigen in a dominant pathogen, activation of dendritic cells with special adjuvants, heterologous boosting reconcile. The present article describes a possible antigen imprinting research algorithm in vaccine preclinical studies. Bibliographical description: Supotnitskiy М. V. The phenomenon of antigenic imprinting infectious epidemic and vaccine-related processes. // Biopreparats (Biopharmaceuticals) 2014; (3) с. 27–39. * * * * * * *
Методами иммунопрофилактики не удается контролировать многие инфекционные болезни, способные к пандемическому распространению (грипп, малярия, ВИЧ/СПИД, лихорадка Денге и др.), что говорит о наличии серьезных пробелов в знаниях об их иммунологии и эпидемиологии. Сложившиеся еще в XIX в. представления о специфическом иммунитете построены на законе Дженнера–Пастера, априори предполагающем, что первый контакт с патогенным микроорганизмом вызывает у организма невосприимчивость (иммунитет), проявляющуюся при повторных контактах с этим же патогеном [1]. Объяснения иммунных ответов на антиген обычно следуют из правила Бернета (Burnet's rule), утверждающего, что антигены рекрутируют специфические лимфоциты из уже существующего репертуара таких клеток и индуцируют их пролиферацию и дифференциацию в эффекторные клетки [2]. Однако в научной литературе накоплены данные о более сложных механизмах взаимодействия иммунной системы человека с патогенными микроорганизмами, чем те, что обычно приводятся в учебных пособиях и руководствах для врачей. Ранее мы рассмотрели в данном контексте феномен антителозависимого усиления инфекции [3]. Цель настоящей работы — обобщить имеющиеся в научной литературе сведения по феномену антигенного импринтингаi в эпидемических, инфекционных и поствакцинальных процессах, и обозначить возможные пути его преодоления при создании новых вакцин. Суть феномена антигенного импринтинга. При повторном контакте иммунной системы с патогенным микроорганизмом или антигенами вакцины, различия между старым вариантом эпитопа антигена, и его новым вариантом, могут быть "незамеченными" иммунной системой примерно так, как в оптический прибор не различаются детали, выходящие за пределы его разрешения. И тогда в процессе антигенной стимуляции первыми активизируются В-клетки памяти, "запомнившие" предыдущий антиген. Далее они дифференцируются в плазмоциты, продуцирующие антитела в отношении этого антигена, хотя иммунная система с ним не контактирует. Образующиеся антитела не способны эффективно нейтрализовать возбудитель инфекционной болезни, выработка же специфических к нему антител тормозится из-за подавления "наивных" В-клеток активизировавшимися В-клетками памяти. Как заметили Kim et al. [7], в данном случае В-клетки памяти формируют "слепое пятно" ("blind spot") иммунной системы. Parsons et al. [8] такой ответ В-клеток памяти назвали замороженным репертуаром (repertoire freeze). Закон Дженнера–Пастера и правило Бернета соблюдаются, но при антигенной дистанции между штаммами (серотипами, серовариантами) возбудителя инфекционной болезни, превышающей размеры "слепого пятна" иммунной системы. Для возбудителей инфекционных болезней, вызывающих феномен антигенного импринтинга, характерны следующие особенности презентации их антигенов иммунной системе инфицированного животного или человека: — наличие эпитопов, экранированных сахарами, и/или с ограниченной иммунодоминантностью; — наличие антигенных детерминант, перекрестно-реагирующих с возбудителями инфекционных болезней, принадлежащих к семействам таксономически малосвязанных патогенов; — выраженная и олигомерная презентация эпитопов иммунной системе; — незначительные различия в аминокислотных последовательностях или в форме антигенных детерминант возбудителя инфекционной болезни и гомологичных белков хозяина; — способность формировать пул длительно живущих В- и Т-клеток памяти; — способность формировать олигоклональный сывороточный профиль после перенесенной инфекции или вакцинации (т. е. происходит ответ только на доминантные эпитопы) [9]. Эпидемиологические и экспериментальные подтверждения феномена антигенного импринтинга. Феномен впервые описан в 1953 г. Davenport et al. [10], обнаружившими низкие титры антител к вирусу серотипа А (H1N1)ii в сыворотке крови людей старше 28 лет и переболевших гриппом до проводимых в начале 1950-х гг. массовых вакцинаций и вакцинированных вакциной на основе вируса этого же серотипа. Однако к вирусу, циркулировавшему ранее, титры антител были неожиданно высокими. Наибольшее количество таких людей приходилось на возрастную группу 35–38 лет, пережившую пандемию гриппа "испанка" в 1918 г. Аналогичные результаты были получены в отношении вируса гриппа серотипа B и его антигенных вариантов [3, 11]. Для объяснения иммунологического феномена Davenport et al. [10] предположили, что во время первого инфицирования вирусом гриппа еще в детском возрасте иммунная система ориентируется на некий доминантный антиген среди циркулирующих штаммов вируса. Последующее экспонирование к вирусам гриппа, антигенно связанным с предыдущим, вызывает подъем уровня антител не на их антигены, а на штамм, вызвавший первую инфекцию. Это наблюдение было кратко резюмировано Francis [4] в виде "доктрины первичного антигенного греха" ("the doctrine of original antigenic sin"). Позже Davenport и Hennessy [12] для определения границ феномена антигенного импринтинга провели вакцинацию моновалентными вакцинами, содержащими инактивированные штаммы различных антигенных вариантов (сероподтипов) вируса гриппа А, циркулировавших среди людей за последние 30 лет. Среди них вирус свиного гриппа (Hsw1N1; swine influenza) — циркулировал во время пандемии "испанки" 1918 г. и некоторое время позже; вирус гриппа А (H0N1)iii — вызывал вспышки гриппа с начала 1930-х гг. до 1943 г.; и вирус гриппа А-prime (H1N1;influenza A-prime) — доминировал в циркуляции среди людей с 1946 г. до начала 1950-х гг. Вакцинация была проведена в группе детей, переболевших гриппом, вызванным вирусом сероподтипа A-prime; в группах взрослых (рекруты), переживших в детском возрасте вспышки гриппа А, и взрослых старше 30 лет. Анализ сывороток показал, что иммунный ответ наиболее выражен к тому сероподтипу вируса гриппа, с которым иммунная система вакцинированного взаимодействовала в детстве. У детей высокие титры антител отмечены в ответ на вакцину на основе вируса гриппа A-prime (H1N1); у рекрутов — на вакцину против вируса гриппа А (H0N1); у людей старше 30 лет — на вакцину на основе вируса свиного гриппа (Hsw1N1). У некоторых волонтеров двух последних групп уже были обнаружены антитела к вирусам гриппа A-prime (H1N1), свидетельствующие о ранее перенесенной инфекции. Реакция человека на введение моновалентных вакцин была типоспецифической. Антитела к вирусу гриппа A-prime, обнаруженные в сыворотке детей, вакцинированных против гриппа A или свиного гриппа, не вступали в перекрестные реакции с вирусами гриппа A или свиного гриппа. Такие же результаты получены в группах рекрутов (антитела к вирусу гриппа A) и людей, старше 30 лет (антитела к вирусу свиного гриппа). К концу 1950-х гг. было окончательно установлено, что антитела к различным сероподтипам вируса гриппа накапливаются в течение всей жизни человека, однако после эпидемических вспышек болезни титр антител бывает наивысшим к тому сероподтипу вируса, который обусловил первое заболевание гриппом в раннем детстве. Francis (1959) обнаружил следующую закономерность распределения антител к разновидностям вируса гриппа типа А среди возрастных групп населения США (табл. 1). Табл. 1. Распределение антител к сероподтипам вируса гриппа типа А в сыворотке людей из разных возрастных групп населения США*
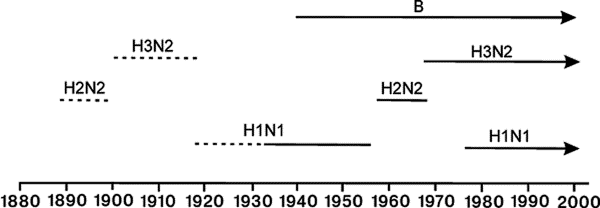
*Цит. по [5]. Классификация сероподтипов вируса гриппа того времени. В конце 1950-х гг. эпидемическая ситуация по гриппу изменилась. Вирусы сероподтипов Hsw1N1, H0N1, H1N1 сменил вирус сероподтипа H2N2 (пандемии азиатского гриппа 1957 и 1959 гг.), затем среди людей появился вирус сероподтипа H3N2 (пандемия Гонконгского гриппа 1968—1970 гг.) (рис. 1).
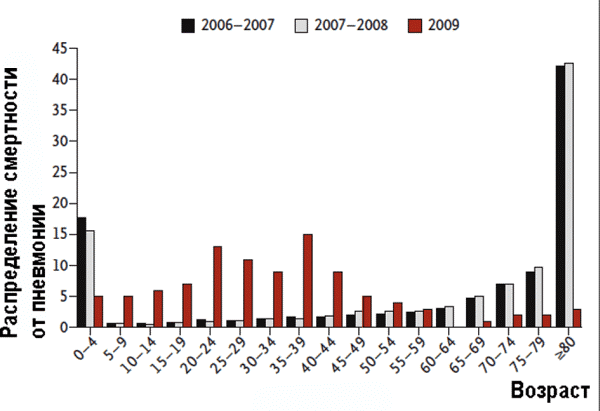
Феномен антигенного импринтинга в 1960-е гг. не только не вызывал сомнения у эпидемиологов и иммунологов, но и использовался ими для разработки методологии археологической серологии. Методология основывалась на определении возрастного распределения антител к различным антигенным вариантам вирусов А и В. Различия в распределении антител среди возрастных групп населения связывали с возникновением анамнестических реакций на вирусы с близкими по структуре антигенами, ранее вызвавшими у человека заболевание гриппом. Благодаря такому подходу было установлено, что вирусы гриппа, сходные с А2N2 и В, циркулировавшие среди людей в начале 1960-х гг., вызывали эпидемии гриппа в 1880–1890-х гг. Для установления истинного сероварианта возбудителя гриппа исследовали сыворотки контингентов однородного возрастного состава (пионерские лагеря, общежития, воинские части) [5]. Появившиеся в циркуляции среди людей вирусы гриппа сероподтипов H2N2 и H3N2 давали собственные анамнестические ответы иммунной системы. Marine и Thomas [14], вакцинируя различные возрастные группы моновалентными инактивированными вакцинами на основе вирусов гриппа серотипа А различных антигенных вариантов (H1N1 и H0N1, H2N2, H3N2), установили, что антигенный импринтинг наблюдается в пределах одного антигенного варианта вируса. Люди, перенесшие первую гриппозную инфекцию, вызванную вирусами сероподтипов H1N1и H0N1, давали анамнестическую реакцию на вакцины, полученные на основе штаммов вирусов этих сероподтипов, но не H2N2 и H3N2, и наоборот. Эффект антигенного импринтинга проявлялся тем интенсивнее, чем больше времени проходило от момента первого контакта иммунной системы с возбудителем гриппа. Masurel и Drescher [15] в опытах на хорьках, последовательно инфицированных с интервалами в три недели разными штаммами вируса гриппа серотипа А (H1N1, Hsw1N1, H0N1, H2N2, H3N2), установили, что вторичное инфицирование может приводить к появлению HCR-антител, т. е. антител с высокой перекрестной активностью (antibodies highly cross-reacting; HCR antibodies) по отношению к штаммам, антигенно тесно связанным по гемагглютинину с теми, что вызвали первый инфекционный процесс. При заражении вирусом гриппа через трехнедельные интервалы антител, специфичных к штамму вируса, вызвавшему первую инфекцию, не появлялось. Однако когда интервал между заражениями увеличивали до 4-5 мес., наблюдался феномен антигенного импринтинга, а HCR-антитела не обнаруживали. Следовательно, образование HCR-антител и антигенный импринтинг — это разные иммунологические феномены. В 1979 г. статистическим анализом заболеваемости населения было обнаружено, что люди, родившиеся до 1956 г., легко перенесли пандемию русского гриппа (1977–1978 гг.). Подавляющее число заболевших приходилось на людей в возрасте до 20 лет, т. е. на ту часть населения, которая не имела контакта с вирусами гриппа серотипа H1N1, вызывавшими вспышки гриппа более 20 лет тому назад. Напротив, лица старше 30 лет составили только 20 % больных, хотя их доля в общей численности населения превышает 50 %, т. е., учитывая низкую заболеваемость в эту эпидемию вообще, люди зрелого и пожилого возраста, имевшие в прошлом контакт с вирусами гриппа H1N1, не болели [16]. Данный феномен наблюдался во всех странах, где велся учет заболевших гриппом, и был объяснен антигенным импринтингом. В экспериментах, выполненных на крысах, установлено отсутствие влияния на анамнестические ответы иммунной системы на вирус сероподтипа H1N1, последующего инфицирования вирусом гриппа других сероподтипов (H2N2, H3N2) [17]. Marine и Thomas [14] подтвердили феномен антигенного импринтинга в масштабном исследовании, выполненном на 687 добровольцах разных возрастов. Добровольцев вакцинировали живыми моновакцинами разных серотипов и изучали анамнестические ответы иммунной системы. В этом же году Couch et al. [18] обнаружили, что после вакцинации инактивированной гриппозной вакциной, полученной на основе штамма вируса A/Scotland/74, в сыворотке 82 % вакцинированных людей присутствовали антитела к вирусу A/HongKong/68, с которым они сталкивались ранее. Только в сыворотке 46 % из них были обнаружены низкие уровни антител к вакцинному штамму A/Scotland/74. Но феномен антигенного импринтинга подтверждался не всегда (см., например, [19]) что требовало объяснения. Границы изменчивости вируса гриппа в пределах его сероподтипов, при которых этот феномен возможен, в 1999 г. смоделировали Smith et al. [20]. По их данным, чем больше антигенное сходство между штаммами вируса гриппа, использованных для вакцинации, и вируса, вызвавшего вспышку гриппа, или антигенов вируса, использованного для повторной вакцинации, тем больше вероятность развития феномена антигенного импринтинга и тяжелого течения болезни у инфицированного пациента. При полной антигенной идентичности вирусов антигенный импринтинг невозможен. Но конкретных величин антигенного различия вирусов, при котором он может возникнуть или быть устраненным, они не привели. Тогда же было обнаружено, что явление антигенного импринтинга может наблюдаться не только при гуморальном, но и при клеточном иммунном ответе на возбудители инфекционных болезней. При повторном реагировании на мутировавшие антигены вируса лимфоцитарного хориоменингита (lymphocytic choriomeningitis virus, LCMV), узнаваемые цитотоксическими Т-клетками, цитотоксический ответ происходил преимущественно в отношении того антигенного варианта вируса, с которым иммунная система человека взаимодействовала первично [21]. В 2010 г. аналогичная роль Т-клеточных ответов иммунной системы человека была описана при лихорадке Денге [22]. Антигенный импринтинг наиболее опасен при развитии повторной инфекции в том случае, если в результате В- и Т-клеточных ответов образуются низкоавидные перекрестно реагирующие антитела на доминирующие антигенные эпипитопы, как, например, это происходит в отношении эпитопов оболочечного белка Е вируса Денге [23]. Такие антитела, образующиеся на ранней стадии повторной инфекции, являются причиной развития другого малоизученного иммунологического феномена — антителозависимого усиления инфекции [24].iv В 1990-х гг. изменилась ситуация в самой эпидемиологии. Началось время глобальных информационных проектов, имеющих целью сверхприбыль от коммерческой реализации вакцин. Представления об эпидемиологии и иммунологии инфекционных болезней, для профилактики которых разработаны вакцины, упростились. Теперь эпидемии уже не были результатом сложных природных и социальных процессов, а возникали из-за "появления нового вируса", что автоматически предполагало новую вакцину, массовую вакцинацию и бюджетное финансирование на ее проведение. Ответы со стороны иммунной системы на вакцину или на возбудителя инфекционной болезни в интересах производителей вакцин стали рассматриваться исключительно как защитные. Феномен антигенного импринтинга исчез из руководств и учебников, и современным российским врачам, в отличие от их советских коллег 1960-х гг., он неизвестен. Описание феномена антитителозависимого усиления инфекции в них и не появлялось. Антигенный импринтинг в пандемию "свиного гриппа" в 2009 г. В 2009 г. Kim et al. [25] подтвердили возможность развития феномена антигенного импринтинга в экспериментах на мышах, используя штаммы A/PR/8/34 (PR8) и A/FM/1/47 (FM1) вируса сероподтипа H1N1. Аминокислотная последовательность HA обоих штаммов была идентична на 92 %. Также они показали, что если проводить последовательную вакцинацию мышей инактивированными вакцинами, полученными на основе разных штаммов вируса гриппа (PR8 и FM1), то при последующем заражении адаптированным штаммом FM1 мыши оказываются менее защищенными от вируса, чем после иммунизации только одним инактивированным FM1. Титр вируса гриппа в легких мышей, вакцинированных сначала PR8, а затем FM1, был в 46 раз выше, чем у мышей, вакцинированных только инактивированным FM1. Мыши, вакцинированные сначала инактивированной вакциной, затем живой, демонстрировали выраженный антигенный импринтинг. Последующее инфицирование животных вирулентным штаммом вируса вызывало у них слабый ответ нейтрализующих антител на этот вирус. Индукция феномена антигенного импринтинга не зависела от введенной дозы вирусов (0,01 или 0,1 LD50) или последовательности, в которой они были введены экспериментальному животному. Choi et al. [26] обнаружили, что 18–20-летние студенты, ранее многократно вакцинированные вакцинами, предназначенными для сезонной вакцинации, реагировали на вакцину против пандемического вируса сероподтипа рH1N1 (pandemic H1N1 2009; pH1N1) значительно слабее, чем ранее не вакцинированные. Однако выяснить, какая вакцинация стала причиной антигенного импринтинга, исследователям не удалось, так как за последние 15 лет в состав вакцин для сезонной вакцинации включалось шесть различных штаммов (!) вируса гриппа сероподтипа H1N1. Было только установлено, что это не комбинированная вакцина, включающая вирус A/Brisbane/59/2007(H1N1), использованная три месяца назад для вакцинации населения. Она не создавала перекрестного защитного эффекта по отношению к вирусу pH1N1. Анализ заболеваемости в разных возрастных группах населения во время глобальной активизации вируса рH1N1 в 2009 г. дал тот же результат, что и подобные анализы заболеваемости, проведенные в начале 1950-х гг. и после пандемии русского гриппа в конце 1970-х гг. У людей, родившихся до 1957 г., антигенный импринтинг стал причиной высоких титров вируснейтрализующих антител, вырабатывающихся как в ответ на вакцинацию, так и на гриппозную инфекцию. В других же возрастных группах антигенный импринтинг повышал смертность заболевших [27–30] (рис. 2).
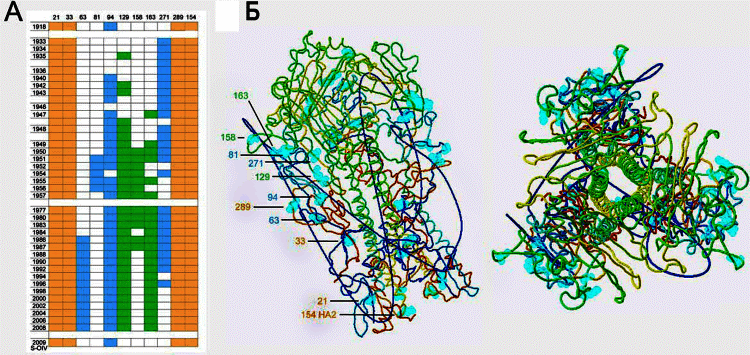
Четыре эпидемиологических исследования распространения вируса пандемического гриппа рH1N1, выполненные в Британской Колумбии (Канада) в 2009 г., обнаружили повышенный риск развития гриппа у лиц, ранее вакцинированных тривалентной инактивированной гриппозной вакциной (trivalent inactivated influenza vaccine, TIV), применяемой для сезонной профилактики гриппа. Авторы связывают его с феноменами антигенного импринтинга, антителозависимого усиления инфекции и с другими еще неизвестными факторами, на необходимость изучения которых они обращают внимание других исследователей [31, 32]. После предшествующих многократных вакцинаций и перенесенных в прошлом заболеваний гриппом, антигенный импринтинг приводит к тому, что в сыворотке крови циркулируют специфические низкоавидные антитела, перекрестно-реагирующие с вирусами гриппа, но не обладающие протективным действием. Например, по данным Monsalvo et al. [33] у умерших пациентов среднего возраста и тех, у кого грипп имел тяжелое течение, специфические низкоавидные антитела (IgG) формировали иммунные комплексы с вирусом, оседавшие в легочной ткани и вызывавшие отек легких, перибронхиолярную мононуклеарную клеточную инфильтрацию и, как результат, гипоксемию. Чем выше был титр таких антигриппозных антител, тем тяжелее протекала болезнь. У пациентов не обнаруживали антител, нейтрализующих pH1N1, и находили вирус гриппа в легочной ткани в высоких титрах. Reichert et al. [30] открыли один из механизмов, благодаря которому антигенная структура вируса гриппа может незначительно меняться, приводя к феномену антигенного импринтинга при повторном взаимодействии с иммунной системой человека. По их данным, HA вируса рH1N1 тесно связан с HA вируса, вызвавшего пандемию гриппа "испанки" в 1918 г., и вирусов, циркулировавших в период с 1930-х по 1943 г. Эволюция вирусов сероподтипа H1N1, циркулировавших в популяциях людей в 1940–1950-е гг. и после его возвращения в 1977 г., происходила через гликозилирование HA (т. е. присоединение остатков сахаров к HA) (рис. 3).
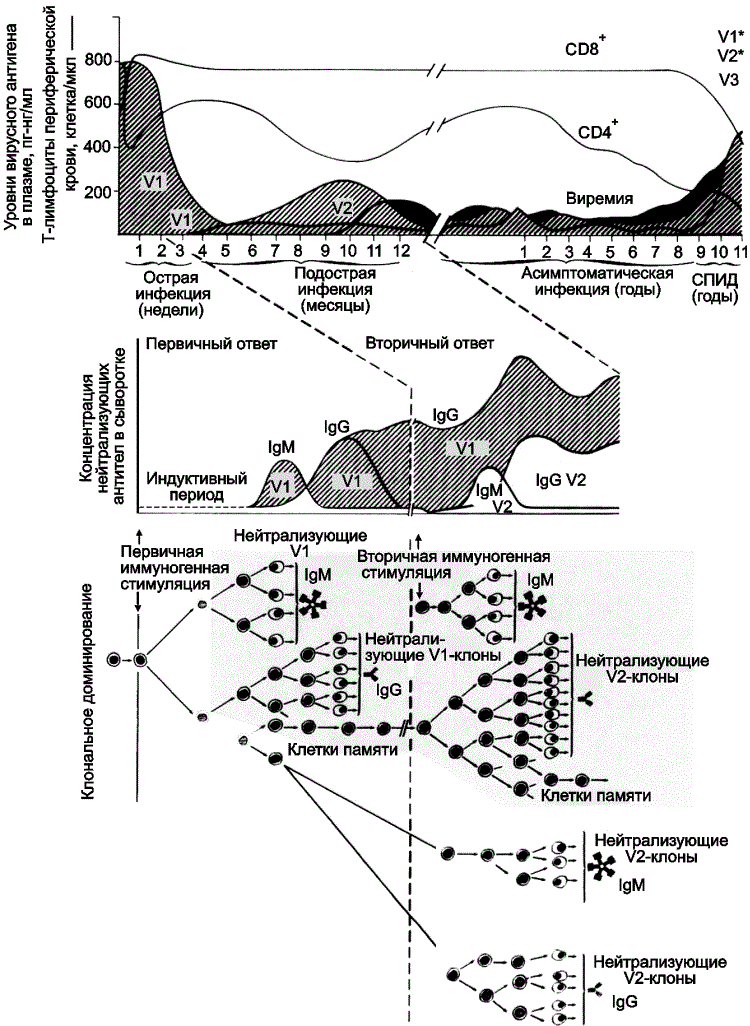
Гликозилирование HA сформировало то антигенное разнообразие среди вирусов гриппа, вызывающих сезонные вспышки болезни, которое проявилось антигенным импринтингом в отдельных возрастных группах после появления в циркуляции вируса pH1N1. Специфичность антигенного импринтинга, проявившаяся защитным эффектом в старших возрастных группах населения, и сравнительные данные по гликозилированию HA вирусов гриппа (см. рис. 3), свидетельствуют в пользу того, что вирус pH1N1 идентичен вирусу, доминировавшему в эпидемиях гриппа первой трети ХХ в. Тогда возникают следующие вопросы: 1) где и как вирус гриппа pH1N1 поддерживался в природе почти 80 лет без гликозилирования HA? 2) каковы механизмы его глобального вовлечения в эпидемические процессы? К сожалению, такие вопросы не интересуют Всемирную организацию здравоохранения, сведшую проблему борьбы с гриппом к приведению массовых вакцинаций. Антигенный импринтинг при ВИЧ-инфекции. Феномен показан при исследовании защитного действия антиВИЧ-вакцин и при инфекционном процессе, вызванном ВИЧ [8, 9, 34]. Первыми на антигенный импринтинг при разработке ВИЧ-вакцин натолкнулись Nara et al. [9]. Они пытались расширить иммунный ответ на антигены ВИЧ в отношении вирусов различного географического происхождения. Введя шимпанзе гликопротеид gp120, полученный из штамма ВИЧ-1 IIIB, и проведя через 175 суток повторную вакцинацию gp120, выделенным из штамма ВИЧ-1 RF, имеющим другое географическое происхождение, они неожиданно для себя обнаружили рост титров антител к gp120 штамма IIIB и отсутствие защитного эффекта при заражении животных ВИЧ-1 RF. Проведенный ими ретроспективный анализ научной литературы показал, что феномен OAS уже был описан для других ретровирусных инфекций, в частности, вызываемых вирусом висны у овец [35] и вирусом инфекционной анемии у лошадей [36] (рис. 4).
При изучении протективного эффекта ВИЧ-вакцины, включающей в качестве антигенного компонента gp120.16, выделенный из ВИЧ-1 SF2, получены сходные результаты. Люди, вакцинированные такой вакциной и имеющие высокие титры антител к gp120.16, оказались восприимчивы к вариантам ВИЧ-1, циркулирующим в их популяции. При развитии ВИЧ-инфекции в их сыворотке крови преобладали антитела к gp120.16 ВИЧ-1 SF2, а не к такому же оболочечному гликопротеину вируса, вызвавшего инфекцию [37]. Феномен антигенного импринтинга обнаружен и при изучении иммунного ответа у ВИЧ-инфицированных пациентов. Выработка антител к ВИЧ у них имеет олигоклональный характер, происходит нарушение соотношения κ/λ типов легких цепей антител, поддерживающееся в течение многих лет независимо от скорости прогрессирования заболевания. Ограниченные (restricted) и при этом стабильно вырабатывающиеся антитела к антигенам ВИЧ у таких пациентов представляют собой одну из причин невозможности выработки плазмоцитам антител к ВИЧ-1, которые бы эффективно связывали сероварианты вируса, образовавшиеся в ходе персистирующего инфекционного процесса [38]. Антигенный импринтинг при малярии. Благодаря работам Pleass et al. [39] удалось показать возможность создания противомалярийной вакцины на основе 19-кДа фрагмента белка MSP119, находящегося на поверхности мерозоитов Plasmodium falciparum — бесполых форм плазмодия. При разрыве эритроцитов мерозоиты попадают в кровь, что приводит к периодическим приступам лихорадки. Связывание специфических антител с белком MSP119 блокирует проникновение возбудителя малярии в эритроциты и активирует его уничтожение фагоцитами. Wipasa et al. [40] в опытах на мышах смоделировали ситуацию гетерогенного ответа на вакцинацию белком MSP119 в популяции людей, живущих в эндемичном по малярии регионе. Ими показано, что заражение мышей P. yoelii YMv вызывает образование антител к нативному MSP119, титр которых после перенесенной мышами экспериментальной малярии исследователи повысили бустерной вакцинацией рекомбинантным белком MSP119. Однако действие, выполненное в обратном порядке, т. е. сначала однократная инъекция (субоптимальная вакцинация) рекомбинантного белка MSP119, а затем инфицирование P. yoelii YM, привело к образованию антител к MSP119, не обладающих протективным действием, и к снижению естественного иммунитета к заражению возбудителем малярии. Антигенный импринтинг при лихорадке Денге. Лихорадка Денге — трансмиссивная болезнь, встречающаяся в странах Южной и Юго-Восточной Азии, Африки, Океании и Карибского бассейна. Обычно имеет мягкое течение и может проходить бессимптомно. Однако в 1–5 % случаев она развивается по типу геморрагической лихорадки (hemorrhagic fever, DHF) с развитием циркуляторного коллапса и с возможностью летального исхода. Причины такого осложнения долго не были ясны. В 1983 г. Halstead et al. [41] обнаружили, что у тайских детей, попавших в клинику в шоковом состоянии после повторного развития у них лихорадки Денге, в сыворотке крови обнаруживаются в основном антитела, специфичные к вирусам серотипов (всего их четыре), вызвавших лихорадку Денге несколько месяцев назад. К серотипам вирусов, обнаруженным у маленьких пациентов вирусологическими методами исследования, антитела образовывались медленно и присутствовали в сыворотке пациентов в низких титрах. Исследователи объяснили данный феномен стимуляцией В-клеток памяти, оставшихся после первого инфицирования, т. е. антигенным импринтингом. Образующиеся в ответ на повторное инфицирование вирусом другого серотипа антитела к вирусам серотипа, вызвавшего первый инфекционный процесс, могут обладать перекрестной специфичностью к штамму вируса, вызвавшего повторное инфицирование пациента, но их авидностьvi невысока. И они не нейтрализуют вирус, вызвавший повторную инфекцию, а наоборот, способствуют его размножению в организме человека, связывая вирусные частицы с Fc-рецепторами (FcR)vii на поверхности макрофагов/моноцитов. По сигнальным путям, инициируемым через Fc-рецептор при взаимодействии с ним комплекса "вирус–Денге-специфическое антитело", запускается экспрессия гена IL-10. Макрофаг начинает продуцировать большие количества IL-10, ингибирующего синтез противовоспалительных цитокинов (IFN-γ, IL 2, 3, 12 и др.) и усиливающего синтез фактора некроза опухолей (TNF), и IL 6. Оба лимфокина повышают проницаемость сосудов. IL-10 также нарушает дифференциацию Т-хелперов на субпопуляции Th1 и Th2, что ведет к нарушению взаимодействия между клеточными и гуморальными звеньями иммунной системы при блокировании размножения вируса Денге. Болезнь развивается в тяжелой клинической форме [3]. Основными антигенами вируса Денге, в отношении которых плазмоцитами синтезируются нейтрализующие антитела, являются оболочечный белок Е и премембранный белок prM. Главную роль в антигенном импринтинге играют эпитопы третьего домена белка Е (ED3). В отношении их происходит выработка антител с широкой перекрестной активностью, но с низкой авидностью [24]. Роль антигенного импринтинга в эпидемических, инфекционных и поствакцинальных процессах. В самом общем виде ее можно представить следующим образом: 1) антигенный импринтинг, развившийся в ответ на инфекционный или вакцинальный процесс, сопровождает человека всю его жизнь и предопределяет реакцию его иммунной системы в инфекционных процессах и структуру заболеваемости населения во время эпидемий (пандемий), вызванных тем же возбудителем инфекционной болезни; 2) при полном антигенном совпадении с возбудителем болезни, сформировавшим В-клетки памяти в прошлом, этими клетками вырабатываются специфические антитела, обладающие протективным действием, развития инфекционного процесса может не произойти. Ретроспективным эпидемиологическим анализом будут обнаружены возрастные группы населения, оказавшиеся мало вовлеченными в эпидемию (пандемию); 3) если между возбудителями инфекционной болезни, вызвавшими первый и последующий (второй) инфекционные процессы, нет антигенного совпадения, но антигенная дистанция между ними настолько мала, что иммунная система не может "отличить" штамм возбудителя инфекционной болезни от того, что сформировал В-клетки памяти во время первого инфекционного процесса, то плазмоциты синтезируют антитела, специфичные к штамму возбудителя инфекционной болезни, распространявшегося в ту пандемию, когда сформировались В-клетки памяти. В результате иммунная система "отрабатывает ложную цель", защитный эффект отсутствует. При ретроспективном эпидемиологическом анализе будут обнаружены возрастные группы населения, понесшие наибольшие потери в данную пандемию; 4) при проявлении антигенного импринтинга в ответ на возбудителя инфекционной болезни или вакцинацию, кроме антител, специфичных к антигену, распознанному иммунной системой человека первым, будут образоваться антитела, реагирующие перекрестно с возбудителями близких по антигенной структуре штаммов, но обладающие по отношению к ним низкой авидностью, и способные усиливать инфекционный процесс (эффект антителозависимого усиления инфекции, см. ниже); 5) если антигенная дистанция между штаммом возбудителя инфекционной болезни, вызвавшим инфекционный процесс в прошлом и вызвавшим новый инфекционный процесс настолько велика, что иммунная система его распознает, то иммунный ответ может быть направлен на противодействие этому штамму. Одновременно сформируются новые В-клетки памяти, которые при последующих вспышках этой же инфекционной болезни будут реагировать с возбудителем болезни, так как описано выше (пп. 1–4); 6) при возможности развития феномена антигенного импринтинга многократная вакцинация и перенесенные инфекционные болезни делают малопредсказуемыми ответы иммунной системы на повторное заражение этими же возбудителями инфекционной болезни: от иммунитета, предотвращающего развитие инфекционной болезни, до ее утяжеления с летальными исходами у заболевших. Поствакцинальные осложнения, связанные с антигенным импринтингом, могут проявляться через десятилетия после ее проведения. Вакцинация одной и той же вакциной может дать противоположные результаты в группах населения, имеющих разную эпидемическую историю и ранее многократно вакцинированных этой же вакциной. Устранение антигенного импринтинга при вакцинацииВ табл. 2 обобщены отдельные подходы, используемые для устранения антигенного импринтинга при вакцинации. Табл. 2 Способы устранения антигенного импринтинга при вакцинации
Судя по датам поступления в редакции научных журналов рукописей статей, работы по созданию нового поколения вакцин, позволяющих "обойти" антигенный импринтинг при массовых вакцинациях населения, ведутся уже не менее 10 лет. Основываются они на знаниях тонких механизмов иммунного ответа применительно к инфекционному процессу, вызванному конкретным возбудителем инфекционной болезни, к сожалению игнорируемых отечественными разработчиками вакцин. Алгоритм исследований антигенного импринтинга при доклиническом изучении вакцин. При планировании таких исследований необходимо учитывать, что при инфекционном процессе и вакцинации иммунная система пациента может реагировать на разные эпитопы даже одних и тех же антигенов. Поэтому выявление феномена антигенного импринтинтинга при доклиническом исследовании вакцин целесообразно вести по двум направлениям: 1) в условиях "после перенесенной инфекционной болезни"; 2) в условиях "после предыдущей вакцинации". Блок-схема алгоритма исследования феномена антигенного импринтинга в доклиническом исследовании вакцины приведена на рис. 5.
Первое направление исследований предполагает заражение адекватных экспериментальных моделей животных возбудителями серотипов, преимущественно циркулирующих в популяциях людей в настоящее время и циркулировавших в прошлом. Такие данные получают сероэпидемиологическими исследованиями. Второе направление предполагает заражение животных штаммами возбудителей инфекционных болезней, распространившихся среди людей уже после вакцинации исследуемыми вакцинами. Общим требованием при доклиническом изучении риска развития антигенного импринтинга у ранее вакцинированных людей должно быть проведение разработчиками вакцин экспериментов, в которых необходимо установить: 1) границы феномена, т. е. очертить "круг" близкородственных видов микроорганизмов (их серотипов или изолятов), при инфицировании которыми вакцинированных людей возможно усиление инфекционного процесса; 2) определить эпитопы антигенов, ответственные за феномен антигенного импринтинга, и эпитопы, ответственные за протективный эффект вакцинации; 3) разработать методические подходы к устранению антигенного импринтинга при вакцинации. Приведенные выше данные показывают, что антигенный импринтинг необходимо учитывать при разработке вакцин и проведении массовых вакцинаций населения против возбудителей инфекционных болезней, содержащих множество поверхностных эпитопов разной перекрестной специфичности, включая антигенно-дрейфующие микроорганизмы, и их более стабильные виды, но циркулирующие конкурентно как множество разных серотипов и штаммов. ЛИТЕРАТУРА1. Мац
АН. Первородный антигенный грех (original antigenic sin) как антипрививочная теологическая
аллюзия. [Интернет]. 2009 [cited 2014 Aug 04]. Available from: http://forums.rusmedserv.com/showthread.php?t=119168. i. В западной научной литературе распространен термин "феномен первичного
антигенного греха" ("phenomenon of original antigenic sin", OAS). Для его
образования Thomas Francis Jr. (1900—1969) [4] использовал библейское выражение "original sin", означающее
"первородный (прародительский, первичный) грех" Адама, отразившийся на всех его потомках.
Из-за произвольных и часто несерьезных толкований термина, связанных с его библейским
происхождением, в данной работе он не используется. В советской научной литературе 1960-х гг. для
описания данного феномена использовался термин "анамнестический ответ" ("anamnestic
response"), см., например, работу Горбуновой [5]. В более поздней работе Ma et al. [6] используется более релевантный термин — "антигенный импринтинг"
("antigenic imprinting"), т. е. "антигенный отпечаток". |
К списку статей В раздел "Прививки" На главную На форум
|